ИЗБРАННЫЕ ГЛАВЫ ИЗ КНИГИ "СОВРЕМЕННАЯ СИЛОВАЯ ТРЕНИРОВКА. ТЕОРИЯ И ПРАКТИКА"
авторы Юрген Хартманн и Харольд Тюннеманн. Издательство
Шпортферлаг, Берлин, 1988 г.![]()
2.2.1. Строение мышцы
Функциональные возможности мускулатуры не могут не интересовать
и представителей большого спорта, и тех, кто занимается спортом
на досуге; и это понятно - в итоге любая спортивная деятельность
совершается при помощи мышц. Значение мускулатуры уже в том, что
на ее долю приходится значительная часть сухой массы тела. Так,
у женщин мышцы составляют 30-35% от общей массы тела, у мужчин
- 42-47%. Силовой тренировкой можно увеличить процентное соотношение
мышц и общей массы тела, а физическое бездействие приводит к уменьшению
мышечной массы и увеличению, как правило, жировой ткани.
2.2.1.1. Виды мышц
Различают гладкие мышцы, скелетные мышцы и сердечную мышцу.
Гладкие мышцы входят в состав внутренних органов, например, в
состав стенок кровеносных сосудов, желудочно-кишечного тракта,
мочевыносящих путей (мочеточник, мочевой пузырь), бронхов. Гладкие
мышцы работают медленно и почти непрерывно, осуществляют относительно
медленные и однообразные движения. Ими нельзя управлять силой
воли. Скелетные мышцы (поперечнополосатые мышцы) удерживают тело
в равновесии и осуществляют движения. Мышцы соединены с костями
при помощи сухожилий. Если мышцы сокращаются, т. е. укорачиваются,
то части скелета через суставы приближаются или удаляются друг
от друга. Работой скелетных мышц можно управлять произвольно.
Они способны очень быстро сокращаться и очень быстро расслабляться.
При интенсивной деятельности они довольно скоро утомляются.
Сердечная мышца по своим функциональным свойствам занимает как
бы промежуточное положение между гладкими и скелетными мышцами.
Так же, как и гладкие мышцы, она практически не поддается воздействию
нашей воли и имеет чрезвычайно высокую сопротивляемость утомлению.
Так же, как и скелетные мышцы, она может быстро сокращаться и
интенсивно работать.
Силовая тренировка эффективно влияет не только на работу скелетных
мышц; благодаря ей изменяется и улучшается функция и состояние
гладкой мускулатуры и сердечной мышцы. Так, например, тренировкой
на силовую выносливость можно увеличить и укрепить сердечную мышцу
и тем самым повысить эффективность ее работы, что в свою очередь
окажет положительное влияние на деятельность скелетных мышц. Хорошо
развитый "мышечный корсет", крепко обхватывающий брюшную полость,
держит внутренние органы, способствует деятельности пищеварительной
системы. Улучшенные функциональные свойства пищеварительной системы,
особенно желудочно-кишечного тракта, а также печени, желчного
пузыря поджелудочной железы, опять же положительно воздействуют
на строение мышц, на энергетическое обеспечение мышечной деятельности
и общее состояние здоровья. Безусловно, силовая тренировка воздействует
на все виды мышечной ткани и на все системы органов человеческого
тела, однако, более подробно мы остановимся на скелетной мускулатуре.
2.2.1.2. Строение скелетной мышцы
Основным элементом скелетной мышцы является мышечная клетка. В
связи с тем, что мышечная клетка по отношению к своему поперечному
сечению) (0,05-0,11мм) относительно длинна (волокна бицепса, непример,
имеют длину до 15 см), ее называют также мышечным волокном. Скелетная
мышца состоит из большого количества этих структурных элементов,
составляющих 85-90% от ее общей массы. Так, например, в состав
бицепса входит более одного миллиона волокон.
Между мышечными волокнами расположена тонкая сеть мелких кровеносных
сосудов (капилляров) и нервов (приблизительно 10% от общей массы
мышцы). От 10 до 50 мышечных волокон соединяются в пучок. Пучки
мышечных волокон и образуют скелетную мышцу. Мышечные волокна,
пучки мышечных волокон и мышцы окутаны соединительной тканью (рис.
1).
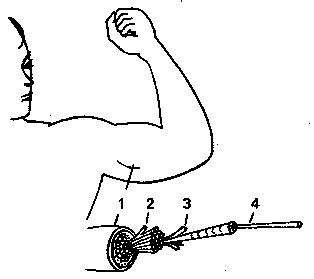
Рис. 1 Схема скелетной мышцы
1 - мышца (5 см), 2 - пучок мышечных волокон (0.5
мм), 3 - мышечное волокно (0,05-0,1 мм), 4 - миофибрилла (0,001-0,003
мм). Цифры в скобках обозначают приблизительный размер поперечного
сечения строительных элементов мышцы.
Мышечные волокна на своих концах переходят в сухожилия. Через
сухожилия, прикрепленные к костям, мышечная сила воздействует
на кости скелета. Сухожилия и другие эластичные элементы мышцы
обладают, кроме того, и упругими свойствами. При высокой и резкой
внутренней нагрузке (сила мышечной тяги) или при сильном и внезапном
внешнем силоном воздействии эластичные элементы мышцы растягиваются
и тем самым смягчают силовые воздействия, распределяя их в течение
более продолжительного промежутка времени. Поэтому после хорошей
разминки в мускулатуре редко происходят разрывы мышечных волокон
и отрывы от костей. Сухожилия обладают значительно большим пределом
прочности на растяжение (около 7 000 N/кв см), чем мышечная ткань
(около 6ОN/кв см), поэтому они гораздо тоньше, чем брюшко мышцы.
В мышечном волокне содержится основное вещество, называемое саркоплазмой.
В саркоплазме находятся митохондрии (30-35% от массы волокна),
в которых протекают процессы обмена веществ и накапливаются вещества,
богатые энергией, например фосфаты, гликоген и жиры. В саркоплазму
погружены тонкие мышечные нити (миофибриллы), лежащие параллельно
длинной оси мышечного волокна. Миофибриллы составляют в совокупности
приблизительно 50% массы волокна, их длина равна длине мышечных
волокон, и они являются, собственно говоря, сократительными элементами
мышцы. Они состоят из небольших, последовательно включаемых элементарных
блоков, именуемых также саркомерами (рис. 2). Так как длина саркомера
в состоянии покоя равна приблизительно лишь 0,0002 мм, то для
того, чтобы, к примеру, образовать цепочки из звеньев миофибрилл
бицепса длиной 10-15 см, необходимо „соединить" огромное
количество саркомеров. Толщина мышечных волокон зависит главным
образом от количества и поперечного сечения миофибрилл.
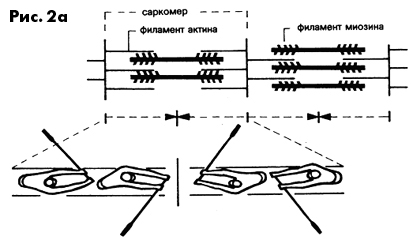
Особенность строения саркомеров позволяет им укорачиваться при
соответствующем нервном импульсе. Процесс сократительного акта
в саркомере можно упрощенно сравнить с движениями гребцов в академической
лодке. Саркомеры состоят из двух видов белковых филаментов: более
тонких - актиновых и более толстых - миозиновых. Из филаментов
миозина с обеих сторон, подобно веслам в лодке, выступают отростки
(миозиновые мостики, см, рис. 2а). Реагируя на нервный сигнал
и последующую химическую реакцию, отростки миозина временно пристыковываются
к филаментам актина (в виде мостиков соединения, а затем отводятся
в „позицию под углом 45"" (см. рис. 2b). За счет этих
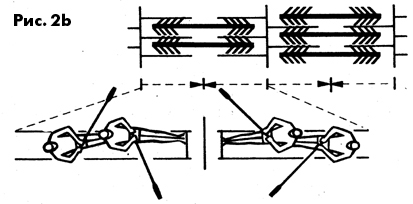
движений, которые можно сравнить с опусканием в воду весел (захват
воды) и последущим гребком, филаменты актина перемещаются между
филаментами миозина. После выполнения „гребка" отростки миозина
примерно так же, как весла, поднимаются из воды, отрываются от
актина и возвращаются в исходное положение. За один такой „гребок"
саркомер укорачивается всего лишь примерно на 1 % своей
длины. Следовательно, для достижения телескопического ' соединения
филаментов, вызывающего эффективное напряжение, требуется большое
число „гребков". Нервная система, выступая в роли „рулевого",
может в зависимости от структуры мышечного волокна и требуемой
величины напряжения, подавать сигналы с частотой от 7 до более
чем 50 „гребков" в секунду. В связи с тем, что огромное количество
саркомеров, расположенных по ходу миофибриллы, включается последовательно,
их единичные минимальные сокращения суммируются, и миофибрилла
сокращается на 25-30%. Так как большое количество миофибрилл расположено
рядом, их относительно небольшие сократительные силы складываются
в суммарную силу мышечного волокна и в итоге в мышцы.
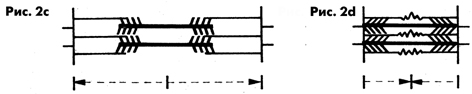
Наиболее благоприятная для образования мостиков длина саркомера
0,0019-0,0022 мм. При этой длине в состоянии покоя филаменты актина
и миозина контактируют настолько удачно, что за единицу времени
может образоваться особенно много мостиковых соединений и тем
самым создаются предпосылки для значительных напряжении в мышце.
При сильном и предельном удлинении мышцы (длина саркомера 0,0024-0,0035
мм) количество контактирующих мостиков уменьшается все больше,
пока отростки миозина перестанут контактировать с филаментами
актина. В результате напряжение в мышце постоянно уменьшается
(рис. 2 с). При сильном и предельном укорачивании мышцы (длина
саркомера 0,0016-0,0013 мм) концы филаментов актина все глубже
проникают между филаментами миозина и их тяга постоянно уменьшается,
образовывать новые мостики становится все сложней. Напряжение
мышцы постоянно спадает (рис. 2 d).
Это явление наблюдается в различных случаях максимального приложения
сил независимо от длины мышцы: и при большой, и при малой длине.
К филаментам актина можно „привязать" лишь относительно небольшое
количество филаментов миозина, поэтому и в начальной, и в конечной
фазе движения большую силу развить нельзя. В средних фазах, в
которых можно навести значительно большее количество мостиков,
силовые возможности увеличиваются (см. рис. 15 и 17). При незначительной
скорости движения мышца имеет возможность создать гораздо большее
количество мостиковых соединений, чем при высокой скорости движения.
При высокой скорости движения, т.е. при высокой скорости сокращения
мышцы, просто-напросто не хватает времени для „стыковки"
филаментов актина и миозина, для одновременного наведения и поддерживания
большого количества мостиков. Поэтому при небольшой скорости мышца
способна на более высокое напряжение и „высвобождает" больше
силы (см. также рис. 9 и 17).

Рис. 3 Механическая модель мышечной деятельности
Мышца: а -при длине покоя, b - в статическом режиме работы,
с - в динамическом режиме работы, d - в растянутом состоянии
СК - сократительный компонент, состоящий из мышечных волокн. или
миофибрилл
Пар - параллельный эластичный компонент, в состав которого входят,
в частности, трубчатые соединительные-тканные оболочки мышечных
волокон (эндомизий) и пучки мышечных волокн (перемизий).
Пос - последовательно включаемый эластичный компонент, образованный.
в частности, сухожилиями.
Внутренняя сила: энергия сокращения (СК) + энергия предварительного
растягивания (Пар + Пос)
Внешняя сила: внешнее сопротивление (оказываемое, например, соперником
или отягощением).
Изложенный процесс сокращения элементарного блока миофибриллы
представляет собой энергетический процесс, в котором химическая
энергия превращается в механическую работу.
Взаимодействие сократительных и эластичных компонентов мышцы наглядно
изображено на механической модели мышцы (рис. 3). Сократительный
компонент мышцы (СК) состоит из миофибрилл. Эластичный компонент
подразделяется на последовательно включаемый эластичный компонент
(Пoc) и параллельно-эластичный компонент (Пар). В состав первого
входят сухожилия и другие элементы соединительной ткани мышцы,
второй образуется, в частности, из соединительно-тканных оболочек
мышечных волокон и их пучков.
Если укорачивается сократительный компонент, то сначала растягивается
Пос (см. рис. 3 b). Лишь после того, как развиваемая в Пос сила
напряжения превысит величину внешней силы (например, сопротивление
соперника или поднимаемого с земли отягощения), сократится вся
мышца. Напряжение Пос во время укорачивания мышцы остается постоянным
(рис. 3 с). Пар. помогает сначала укоротить сократительный компонент,
а затем вернуть его к длине покоя. Если мышца растягивается, то
внешняя сила настолько сильно удлиняет Пос, что в конце концов
за ним приходится следовать и сократительному компоненту (СК)
(см. рис. 3 d). При длине покоя мышца может развить очень высокое
напряжение. Во-первых, потому что оптимальная степень контакта
филаментов актина и миозина позволяет создать максимальное количество
мостиковых соединений и тем самым активно и сильно развить напряжение
сократительного комнонента. Во-вторых, потому что эластичный компонент
мышцы уже как пружина предварительно растянут, уже создано дополнительное
напряжение. Активно развитое напряжение сократительного компонента
суммируется с упругим напряжением, накопленным в эластичном компоненте,
и реализуется в одно высокое, результирующее напряжение мышцы
(см. 2.7.). Последующее предварительное растяжение мышцы, которое
значительно превосходит состояние при длине покоя, приводит к
недостаточному контакту филаментов актина и миозина. При этом
заметно ухудшаются условия для развития значительного и активного
напряжения саркомеров. Тем не менее при большом предварительном
растягивании задействованных мышц, например, при широком замахе
в метании копья, спортсмены достигают более высоких результатов,
чем без замаха. Этот феномен объясняется тем, что увеличение предварительного
напряжения 'эластичного компонента превосходит снижение активного
развития напряжения сократительного компонента.
За счет целенаправленной силовой тренировки (метод многократной
субмаксимальной нагрузки, см. 9.2.1.1.) увеличивается поперечное
сечение и количество как сократительных элементов, миофибрилл,
так и других соединительно-тканных элементов мышечного волокна
(митохондрии, фосфатные и гликогенные депо и т. д.). Правда, этот
процесс приводит к прямому увеличению сократительной силы мышечных
волокон, а не к немедленному увеличению их поперечного сечения.
Лишь после того, как это развитие достигнет определенного уровня,
продолжение тренировок по развитию силы может способствовать увеличению
толщины мышечных волокон и тем самым увеличению поперечного сечения
мышцы (гипертрофия).
Таким образом, увеличение поперечного сечения мышцы происходит
за счет утолщения волокон (увеличение саркомеров в поперечном
сечении мышцы), а не за счет увеличения числа мышечных.волокон,
как часто ошибочно предполагают. Количество волокон в каждой отдельно
взятой мышце обусловлено генетически и, как показывают научные
исследования, это количество нельзя изменить при помощи силовой
тренировки. Интересно, что люди значительно отличаются по количеству
мышечных волокон в мышце. Спортсмен, в бицепсе которого содержится
большое количество волокон (см. рис. 60 а), имеет лучшие предпосылки
увеличить поперечное сечение этой мышцы тренировкой, направленной
на утолщение волокон, чем спортсмен, бицепс которого состоит из
относительно небольшого количества волокон. У наиболее способных
представителей видов спорта, требующих максимальной и скоростной
силы, при планомерной и настойчивой тренировке доля мышц к общей
массе тела увеличивается до 60% и более процентов (рис. 4). Сила
скелетной мышцы, как уже отмечалось, зависит главным образом от
ее поперечного сечения, т. е. от количества и толщины миофибрилл,
параллельно расположенных в волокнах, и складывающегося из этого
количества возможных мостиковых соединений между филаментами миозина
и актина.
Таким образом, если спортсмен увеличивает поперечник мышечных
волокон, то он увеличивает и свою силу. Однако сила и мышечная
масса увеличиваются не в одинаковой мере. Если мышечная масса
увеличивается в два раза, то сила увеличивается. примерно, в три
раза. У женщин сила составляет 60-100 N/кв см (6-10 кг/кв см^,
ay мужчин - 70-120 N/кв см. Большой разброс этих показателей (отдача
силы на 1 кв см площади поперечного сечения) объясняется разными
факторами, как зависящими, так и не зависящими от тренировки,
например, внутримышечной и межмышечной координацией, энергетическими
запасами и строением волокна.
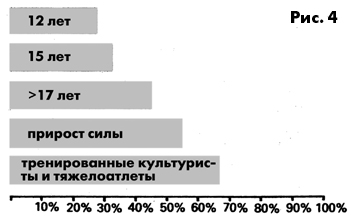
Рис. 4 Процентная доля мышечной массы к общей массе тела у
лиц мужского пола, занимающихся спортом (у супертяжеловесов
доля мышц обычно на 10-20% меньше)
2.2.1.3. Типы волокон скелетной мышцы
Каждая мышца состоит из волокон, обозначаемых как ST-волокна
(slow twitch fibres) - медленно сокращающиеся и FT-волокна - быстро
сокращающиеся (fast twitch fibres). ST-волокна, обладающие высоким
содержанием миоглобина (красный мышечный пигмент), называют также
красными волокнами. Они включаются при нагрузках в пределах 20-25%
от максимальной силы и отличаются хорошей выносливостью. FT-волокна,
обладающие по сравнению с красными волокнами небольшим содержанием
миоглобина, называют также белыми волокнами. Они характеризуются
высокой сократительной скоростью и возможностью развивать большую
силу. По сравнению с медленными волокнами они могут вдвое быстрее
сокращаться и развить в 10 раз большую силу (см табл. 1). FT-волокна,
в свою очередь, подразделяются на FTO-и FTG-волокна: наименование
их определяется способом получения энергии. Получение энергии
в FTO-волокнах происходит так же, как и в ST-волокнах, преимущественно
путем окисления, в результате чего глюкоза и жиры в присутствии
кислорода разлагаются на двуокись углерода (СО2 и воду (Н20).
В связи с тем. что этот процесс разложения протекает относительно
экономично (на каждую молекулу глюкозы при разложении мышечного
гликогена для получения энергии накапливается 39 энергетических
фосфатных соединений), FTO-волокна имеют также относительно высокую
сопротивляемость утомляемости (см. 2.2.3.). Накопление энергии
в FTG-волокнах происходит преимущественно путем гликолиза, т.
е. глюкоза в отсутствии кислорода распадается до еще относительно
богатого энергией лактата. В связи с тем, что этот процесс распада
неэкономичен (на каждую молекулу глюкозы для получения энергии
накапливается всего лишь 3 энергетических фосфатных соединения),
FTG-волокна относительно быстро утомляются, но тем не менее они
способны развить большую силу и, как правило, включаются при субмаксимальных
и максимальных мышечных сокращениях (см. 9.2.).
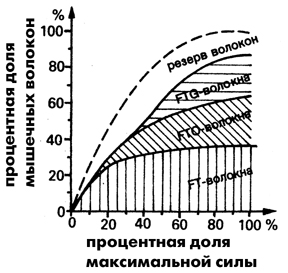
Рис. 5 Включение различных типов мышечных волокон в движение
при увеличивающихся сопротивлениях (преобразовано по Костилу)
Процесс взаимодействия различных типов волокон до настоящего времени
выяснен не до конца. Схематично он может быть изложен так.
При нагрузках менее 25% от максимальной силы сначала начинают
функционировать преимущественно медленные волокна. Как только
их запасы энергии иссякают, „подключаются" быстрые волокна.
После того, как израсходуются энергетические запасы быстрых волокон,
работу придется прекратить, наступает истощение. Если же силовая
нагрузка возрастает от низких до максимальных величин, то согласно
Костиллу (1980 г.) возникает так называемый „эффект рампы",
когда почти все волокна вовлекаются в движение (рис. 5). Изображенный
на рис. 5 принцип участия различных типов волокон в мышечной деятельности
действителен, по всей вероятности. для всех движений. Сначала
включаются медленные волокна, а чуть позже, когда потребность
в силе превысит 25% от максимальной, в активность вступают и быстрые
волокна.
При взрывных движениях временной промежуток между началом сокращения
медленных и быстрых волокон минимален (всего несколько мс). Таким
образом, начало сокращении у обоих типов волокон происходит почти
одновременно, однако быстрые волокна укорачиваются значительно
быстрее и раньше достигают своего силового максимума (приблизительно
за 40-90 мс), чем медленные волокна (приблизительно за 90-140
мс), поэтому за взрывную силу, которая должна быть реализована
в течение 50-120 мс, „отвечают" главным образом быстрые волокна
(см. рис. 51 а).
Скорость сокращения быстрых и, хотя в значительно меньшей степени,
медленных волокон можно повысить тренировкой по специальным методам
развития максимальной и скоростной силы (см. 9.2.1.3. и 9.2.2.).
Упражнения на многократное взрывное преодоление субмаксимальных
сопротивлений могут помочь, например, уменьшить время сокращения
(от начала сокращения до достижения силового максимума) быстрых
волокон приблизительно до 30 мс и медленных волокон примерно до
80 мс.
Наименование „быстрое волокно" или „медленное волокно"
вовсе не означает, как иногда ошибочно истолковывают, что относительно
быстрые движения реализуются исключительно быстрыми волокнами,
а медленные движения лишь медленными волокнами. Для включения
волокон в работу решающее значение имеет мобилизуемая сила, т.
е. величина, требуемая для передвижения массы (веса), а также
величина ускорения этой массы.
В соответствии с имеющейся на сегодняшний день информацией и большое
ускорение незначительного веса (большая скорость движения), и
незначительное ускорение большого веса (медленная скорость движения)
осуществляется за счет интенсивного участия быстрых мышечных волокон.
Взрывные силы, направленные на преодоление неподвижных сопротивлений
(статический режим работы, скорость движения = 0 м/с), также вызываются
прежде всего быстрыми волокнами. Каждый человек обладает индивидуальным
набором ST-и FT-волокон, количество которых, как показывают научные
исследования, нельзя изменить при помощи специальной тренировки.
В среднем человек имеет примерно 40% медленных и 60 % быстрых
волокон. Но это средняя величина (по всей скелетной мускулатуре),
мышцы же выполняют различные функции и поэтому могут значительно
отличаться друг от друга составом волокон. Так, например, мышцы,
выполняющие большую статическую работу (камбаловидная мышца),
часто обладают большим количеством медленных ST-волокон, а мышцы,
совершающие преимущественно динамические движения (бицепс), имеют
большое количество FТ-волокон. Однако как показывают многочисленные
исследования, встречаются и значительные индивидуальные отклонения.
У бегунов на длинные дистанции в икроножной мышце и пловцов-стайеров
в дельтовидной мышце было обнаружено. 90% медленных волокон, а
у спринтеров в икроножной мышце до 90% быстрых волокон. Эти индивидуальные
поразительные величины распределения волокон, вероятно, нельзя
объяснить тренировкой, - они обусловлены генетически. Это подтверждается,
в частности, тем, что, несмотря на гармоничное развитие скоростной
силы рук и ног, боксер или фехтовальщик может, например, иметь
чрезвычайно „быстрые ноги" и „медленные руки". Прирожденное
количество быстрых FT-волокон является, видимо, причиной этого
несоответствия. Тот факт, что у хороших представителей видов спорта,
где особенно требуется выносливость (марафонцы, велосипедисты-шоссейники
и т. и.), в основном преобладают медленные ST-волокна, а высококлассные
атлеты, которые демонстрируют скоростную силу (спринтеры, копьеметатели,
толкатели ядра), обладают высоким процентом быстрых FТ-волокон,
свидетельствует об особом предрасположении именно к этим видам
спорта. На первый взгляд кажется, что это положение спорно, так
как у тяжелоатлетов - победителей различных соревнований - обнаружено
чрезвычайно уравновешенное соотношение FT-и ST-волокон. Однако
следует учитывать специфическую работу тяжелоатлета: опору и удержание,
которая в значительной степени выполняется посредством ST-волокон.
Таблица 1. Упрощенное изображение спектра мышечных волокон
| Характеристика (функция) | Тип волокна | ||
| FT-волокна FTG-волокна |
FTO-волокна | ST-волокна | |
| Физиологическая характеристика: | |||
| - скорость сокращения | быстрая | быстрая | медленная |
| - сила сокращения | очень высокая | высокая | незначительная |
| - реакционная способность. | быстрая | быстрая | медленная |
| - аэробная выносливость | плохая | хорошая | очень хорошая |
| Биохимическая характеристика: | |||
| - накопление энергии | гликогенное | гликогенное/окислительное | окислительное |
| отложения фосфата | +++ | ++ | + |
| - отложения гликогена | +++ | ++(+) | ++ |
| - жировые отложения | + | +(+) | ++(+) |
| - содержание митохондрий | + | ++ | +++ |
| - капилляризация | незначительная | хорошая до очень хорошей | очень хорошая |
| Функция: | нагрузки в субмаксимальной зоне, проявление максимальной и скоростной силы |
выносливость и силовая выносливость, статическая работа на опору и удержание |
|
| +++ - значительные, ++ - средние, + - незначительные |
|||
Соответствующей силовой тренировкой можно относительно быстро
преобразовывать FT-волокна в FTO-волокна. Это дает возможность
достигать хорошей выносливости даже тем спортсменам, которые,
имея много быстрых FT-волокон, казалось бы более подходят для
проявления максимальной и скоростной силы.
Несмотря на то, что тренировкой нельзя изменить унаследованное
соотношение между ST- и FT-волокнами, свойства волокон, хоть и
в определенных пределах, все же приспосабливаются к предъявляемым
специфическим раздражениям (поперечное сечение, время сокращения,
оснащение энергоносителями и митохондриями и т. д.).
2.2.2. Нервно-мышечные процессы
2.2.2.1. Двигательная единица
Отросток двигательного нерва, находящегося в спинном мозге (двигательная
клетка переднего рога), достигает мышечного волокна. Нервная клетка
иннервирует своими отростками большое количество мышечных волокон.
Нервная клетка и связанные с ней мышечные волокна называются двигательной
единицей (рис. 6).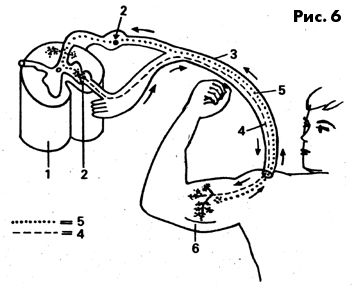
Рис. 6 Схема двигательной единицы
1 - костный мозг, 2 - тело клетки. 3 - нерв спинного мозга, 4
- двигательное нервное волокно, 5 - чувствителыюе нервное волокно,
6 -мышца с мышечными волокнами.
Состав мышц может очень различаться по количеству двигательных
единиц, а двигательные единицы в свою очередь могут состоять из
самого различного количества мышечных волокон. Все мышечные волокна
одной двигательной единицы относятся к одному и тому же типу волокон
(FT- или ST-волокна). Мышцы, в функцию которых входит выполнение
очень тонких и точных движений (например, мышцы глаз или пальцев
руки, обладают обычно большим количеством двигательных единиц
(от 1500 до 3000); в их состав входит небольшое количество мышечных
волокон (от 8 до 50). Мышцы, выполняющие относительно грубые движения
(например, большие мышцы конечностей), обладают, как правило,
значительно меньшим количеством двигательных единиц, но с большим
числом волокон на каждую (от 600 до 2000). Так, например, бицепс
(см. рис. 60 а) может содержать в своем составе более миллиона
волокон. Эти мышечные волокна вместе со своими нервными окончаниями
образуют более чем 600 двигательные единиц, так что одна двигательная
клетка переднего рога спинного мозга иннервирует своими отростками
около 1500 мышечных волокон. В большеберцовой мышце около 1600
и в мышцах спины до 2000 мышечных волокон иннервируются одной
клеткой переднего рога, образуя таким способом в каждом случае
двигательную единицу. Однако количество волокон в двигательных
единицах какой-либо мышцы не одинаково, например, в бицепсе может
быть 1000, 1200, 1400 или 1600 волокон.
Принадлежность мышечных волокон к определенной двигательной единице
задается от природы и не может быть изменена тренировкой.
Двигательные единицы активизируются по закону „все или ничего".
Таким образом, если от тела двигательной клетки переднего рога
спинного мозга посылается по нервным путям импульс, то на него
реагируют или все мышечные волокна двигательной единицы, или ни
одного. Для бицепса это означает следующее: при нервном импульсе
необходимой силы укорачиваются все сократительные элементы (миофибриллы)
всех (примерно 1500) мышечных волокон соответствующей двигательной
единицы.
Сила двигательной единицы зависит, в частности, и от количества
ее мышечных волокон. Двигательные единицы с небольшим количеством
волокон при единичном сокращении развивают силу тяги всего лишь
в несколько миллиньютонов. Двигательные единицы с большим количеством
волокон - несколько ньютонов. Силовой потенциал отдельной двигательной
единицы относительно небольшой, поэтому для выполнения движения
одновременно „подключается" несколько двигательные единиц.
Чем выше преодолеваемое сопротивление, тем больше двигательных
единиц должно выполнять движение.
Каждая двигательная единица имеет свой индивидуальный порог возбуждения,
который может быть низким или высоким. Если импульсный залп (раздражение
нерва, вызывающее сокращение мышцы) слаб, то тогда активизируются
лишь двигательные единицы, обладающие низким порогом возбуждения.
Если импульсный залп усиливается, начинают реагировать дополнительные
двигательные единицы с более высоким порогом возбуждения.
С увеличением сопротивления активизируется все больше двигательных
единиц. Быстрота индивидуальных порогов возбуждения зависит главным
образом от состояния двигательных единиц. Для продолжения деятельности
двигательных единиц, которые утомляются от: а) накопления
кислых продуктов обмена веществ (лактат, СО2); б) истощения энергоносителей
(энергетические фосфаты, гликоген и т.п.); в) нервного перевозбуждения
(в двигательной единице или в коре головного мозга), требуется
все больше и больше волевых усилий.
2.2.2.2. Внутримышечная координация и частота импульсов
Постоянное изменение количества участвующих в движении двигательных
единиц (пространственная суммация) и изменение частоты нервных
импульсов (временная суммация) регулируется очень тонкой градацией
сократительной силы мышцы.
Пространственная суммация. Для выполнения движения может
быть задействовано различное количество двигательных единиц благодаря
механизму ступенчатого развития силы. Однако этот механизм, обусловленный
дифференцированным строением мышц, очень неоднороден. Количество
ступеней определяется количеством двигательных единиц, из которых
состоит мышца; размер ступеней зависит, в частности, от количества,
поперечника и строения мышечных волокон, которыми располагает
соответствующая двигательная единица. Например, в состав мышц
пальцев кисти входит чрезвычайно много двигательных единиц с небольшим
количеством волокон (многочисленные маленькие ступени), поэтому
сила, с которой они выполняют движения, может быть „градуирована"
при помощи пространственной суммации гораздо тоньше, чем сила
бицепса, обладающего относительно малым количеством двигательных
единиц и большим числом волокон (немногочисленные большие ступени).
Временная суммация. Если двигательная единица активизируется
лишь путем искусственного раздражения, например, электрической
стимуляцией, то все ее мышечные волокна укорачиваются, а затем
снова расслабляются (рис. 7а).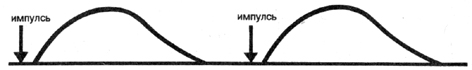
Рис. 7а Схема феномена временной суммации
Однако в здоровом организме в естественных условиях произвольные
одиночные импульсы или сокращения не возникают. Сокращение мышцы
всегда обуславливается действием серии импульсов в секунду. Если
второй сократительный импульс подаётся еще до окончания фазы расслабления
волокон, то в этом случае второе сокращение наслоится на первое.
Следствие этого - более высокое развитие силы. Если нужно развить
большую силу, то второй импульс уже должен достичь волокон двигательной
единицы незадолго до окончания фазы сокращения. Тогда волокна
снова сократятся еще до начала фазы расслабления; спад напряжения
или силы в этом случае невозможен (рис. 7 b). Последующие сокращения
вытекают из предыдущих. 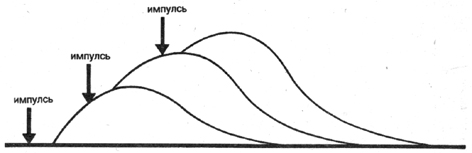
Рис. 7Ь Схема феномена временной суммации
Когда, наконец, многочисленные нервные импульсы начинают следовать
друг за другом достаточно быстро, отдельные сокращения полностью
перекрываются. Таким способом, в отличие от одиночного сокращения,
достигаются гораздо более сильные сокращения мышечных волокон,
что приводит к 3-4-х кратному увеличению силы. Это явление называется
титаническим сокращением (рис. 7 с). Необходимая для полного тетанического
сокращения частота импульсов определяется соответствующим типом
волокна двигательной единицы. В связи с тем, что быстрые FT-волокна,
по сравнению с медленными ST-волокнами, гораздо быстрее сокращаются
и расслабляются, импульсы также должны в более короткие промежутки
попадать в волокна для того, чтобы можно было помешать их расслаблению
и тем самым развить большую силу.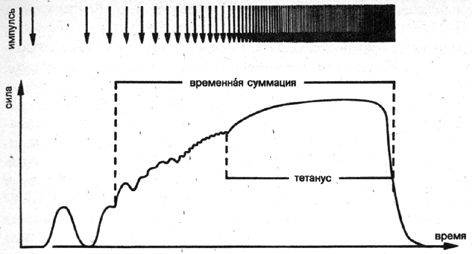
Рис. 7с Схема наслаивания отдельных сокращений до момента возникновения
тетанического сокращения рутем увеличения частоты импульсов
Поэтому у быстрых двигательных единиц импульсы низкой частоты
(7-10 за с) вызывают лишь незначительное напряжение и такую же
силу, импульсы средней частоты (25-30 за с) соответственно умеренное
напряжение и силу, импульсы высокой частоты (от 45 за с и выше)
- максимальное напряжение и максимальную силу (рис. 8). 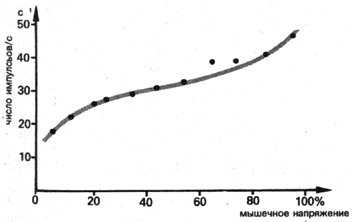
Рис. 8 Взаимосвязь между числом нервных импульсов в секунду
и силой сокращения мышц в процентах к максимальной силе; среднее
значение многочисленных двигательных единиц; отведение от абдуктора
мизинца (по Бигланду и Липпольду)
Для медленных двигательных единиц, состоящих из ST-волокон, уже
20 импульсов в секунду может быть достаточно для исчерпания их
силового потенциала. Лишь при одном, самом благоприятном для соответствующей
двигательной единицы, временном промежутке между импульсами можно
добиться оптимального эффекта временной суммации. Более высокая
частота импульсов для данной двигательной единицы не может вызвать
более сильного сокращения и, следовательно, увеличения силы. Продолжительность
титанического сокращения может превышать продолжительность одиночного
сокращения в десятки и тысячи раз.
Мышца, состоящая большей частью из ST-волокон, более устойчивых
к воздействию утомляемости, может поддерживать титаническое сокращение
обычно значительно дольше, чем мышца, в составе которой содержатся
преимущественно быстро утомляющиеся FT-волокна. В упрощенном изложении
„кооперирование" пространственной и временной суммации происходит
следующим образом: небольшие силовые потребности удовлетворяются
медленными, состоящими из ST-волокон двигательными единицами,
обладающими низким порогом возбуждения. При увеличении силовых
потребностей включаются двигательные единицы, имеющие более высокий
порог возбуждения (пространственная суммация). Одновременно за
счет повышения частоты импульсов увеличивается силовая отдача
уже работающих низкопороговых единиц (временная суммация). При
дальнейшем увеличении силовых потребностей в работу постепенно
будет включаться все больше и больше быстрых двигательных единиц,
которые могут „стартовать" с более высоких частот и вовлекать
в активное состояние больший диапазон частот. Для преодоления
максимальных сопротивлений подготовленные в силовом отношении
спортсмены включают около 85% своих двигательных единиц с оптимальными
импульсными частотами. В связи с тем, что „медленные" единицы
имеют меньше мышечных волокон и по этой причине развивают меньше
силы, чем „быстрые" единицы, часто уже при усилии в 25% мобилизуется
около 50% имеющихся в распоряжении единиц.
Участие относительно большого количества малых двигательных единиц
в незначительной силовой работе позволяет проводить более тонкую
регуляцию мышечной деятельности, чем при высоких силовых нагрузках.
Процессы временной суммации (частота импульсов) согласно результатам
последних исследований условно поддаются тренировке, пусть даже
эта тренировка проводится в очень сложных общих взаимосвязях.
Тренированная двигательная единица может быстрее укорачиваться
(см. 2.2.1.3.), обрабатывать" более высокие импульсные частоты
и развивать большую силу.
Когда скоростная сила, которая реализуется главным образом быстрыми
FT-волокнами, противодействует умеренным и высоким сопротивлениям,
происходит активизация большого количества двигательных единиц
с короткой серией импульсов. Эта, так называемая, стартовая
иннервация вызывает нарастающий и сильный процесс сокращения.
За взрывным началом сокращения следует сигнальная блокировка (биоэлектрическое
молчание), во время которой двигательные единицы сокращаются с
высокой скоростью. Такие скоростно-силовые движения называются
также баллистическими движениями. Они заранее программируются
в головном мозге и осуществляются с такой высокой скоростью, что
во время их выполнения обратная связь не срабатывает, в результате
чего движение невозможно исправить в ходе его выполнения. Продолжительность
биоэлектрического молчания, следующего за стартовой иннервацией,
зависит главным образом от величины преодолеваемого сопротивления.
Если сопротивление настолько велико, что ускорения при свободном
сокращении больше не происходит, то следует новая серия импульсов,
сопровождаемая биоэлектрическим молчанием, благодаря которой обеспечивается
дальнейшее ускорение. Если же сопротивление настолько велико,
что серия импульсов и последующая сигнальная блокировка не появляются,
то сопротивление будет преодолеваться импульсами очень высокой
частоты. Движения, которые характеризуются короткой серией (сериями)
импульсов с последующей сигнальной блокировкой и баллистическим
сокращением, имеют резко выраженный скоростно-силовой характер.
Движения, характеризуемые рядом импульсов очень высокой частоты,
имеют характер максимальной силы.
Когда скелетная мышца работает на силовую выносливость и преодолевает
легкие или умеренные сопротивления, при которых частота импульсов
не достигает максимума, деятельность двигательных единиц осуществляется
попеременно (асинхронная деятельность).
Это означает, что в соответствии с требуемым усилием активизируется
лишь определенная часть двигательных единиц и таким образом происходит
движение. Другая часть двигательных единиц находится в неактивном
состоянии и укорачивается пассивно. При возрастании утомляемости
двигательные единицы, бывшие до сего времени активными, выключаются,
а вместо них начинают активно работать другие, неактивные до сих
пор, двигательные единицы.
В обычных условиях человек, выполняя статическую или динамическую
работу преодолевающего характера (см. 2.3.), не может одновременно
включать в движение все двигательные единицы мышцы. Высокотренированные
атлеты тех видов спорта, в которых сила является основным компонентом
результативности (тяжелая атлетика, борьба, легкоатлетические
метания), для выполнения движения способны активно и одновременно
подключать до 85% своих мышечных волокон и тем самым развивать
большую силу. Нетренированные лица могут обычно активизировать
лишь до 60%. Умение управлять двигательными единицами синхронно
называется внутримышечной (интрамышечной) координацией. Ее
уровень можно считать высоким, если спортсмен, с одной стороны,
обладает ярко выраженной способностью дифференцировать силу и,
с другой стороны, может одновременно активизировать высокий процент
двигательных единиц. Под воздействием гипноза или при электрической
стимуляции (100 гц и выше) нетренированный человек может одновременно
задействовать значительно больше двигательных единиц и тем самым
увеличить свою силу почти на 35%.
Тренированный человек при независящих от усилия воли условиях
может увеличить свой силовой потенциал лишь на 10%. Разница между
произвольно мобилизованной максимальной силой и непроизвольно
активизированной силой называется дефицитом силы. В тренировочной
практике дефицит силы определяется чаще всего разницей в силе,
развиваемой в статическом и динамически-уступающем режимах (см.
2.3.). Такое определение возможно потому, что сила, развиваемая
при принудительном растягивании мышц (динамическая работа уступающего
характера) обычно на 10-35% превышает силу, которая может быть
мобилизована при статическом режиме работы. Таким образом, в показателях
силы, достигаемых, с одной стороны, электрической стимуляцией
мышц в статическом режиме и, с другой стороны, принудительным
растягиванием мышц в динамическом режиме, имеется полное соответствие.
При уступающем режиме работы независимо от воли подключаются дополнительные
двигательные единицы, т.е. в этих условиях величина силы практически
не зависит от уровня внутримышечной координации. При этом необходимо
иметь в виду, что вызванную силу и произвольную можно сопоставлять
друг с другом лишь тогда, когда они прикладываются в сопоставимых
условиях (например, при одном и том же угле в суставах).
Экспериментально удалось доказать, что величина развития силы
при принудительном растягивании мышц увеличивается вместе с увеличением
скорости, в то время как при преодолевающем режиме работы она
при увеличении скорости снижается (см. 2.2.1.2. и рис. 9).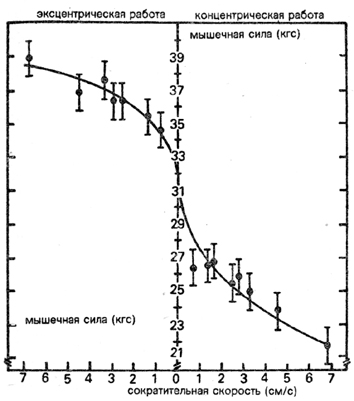
Рис. 9 Скоростно-силовая пропорция у одной из мышц-сгибателей
предплечья при выполнении эксцентрической и концентрической работы
(по Коми)
Итак, при увеличении скорости сокращения разница между возможной
мобилизацией силы при преодолевающем и уступающем режимах работы
продолжает расти.
Однако, нужно сказать, что это утверждение только затрагивает
детей в возрасте до 14 лет. Разница в силе, которую развивают
дети этого возраста при преодолевающем и уступающем режимах работы,
очень незначительна.
Таким образом, чем больше у взрослых различий в показателях силы
при работе в статическом и динамически-уступающем режимах, тем
меньше их максимальная сила по сравнению с абсолютной, тем больше
их дефицит силы и тем ниже Уровень их внутримышечной координации.
В этом случае мышечную силу можно развить при помощи тренировки,
направленной на одновременное включение в движение возможно большего
количества двигательных единиц (метод кратковременных максимальных
напряжений, см. 9.2.1.3.). Сила отдельного мышечного волокна или
отдельной двигательной единицы вряд ли будет увеличена таким путем,
т.е. мышечный поперечник (и вес тела спортсмена) или совсем не
увеличится, или увеличится незначительно. Если разница в силе
при статическом и динамически-уступающем режимах работы относительно
невелика, то резервов для развития силы путем улучшения внутримышечной
координации практически нет. Силу нужно развивать другим путем,
например, увеличивая поперечное сечение мышцы. Дефицит силы представляет
собой своеобразный показатель имеющегося потенциала развития максимальной
силы без увеличения поперечника мышцы и тем самым - веса тела.
Различная способность у нетренированных людей и высококвалифицированных
спортсменов к мобилизации максимально большого количества двигательных
единиц показывает, что подготовленный в силовом отношении человек
обычно имеет не только большую мышечную массу, но и более обширную
возможность ее использования - до 85%. Это также означает, что
возможность развития силы совершенствованием внутримышечной координации
у подготовленных высококвалифицированных спортсменов в сравнении
с неподготовленными ограничена. По этой причине увеличение поперечника
мышцы является самым основным методом для повышения базового потенциала
силы.
2.2.2.3. Межмышечная координация
В спорте практически нельзя выполнять движения с использованием
какой-либо одной мышцы. Для решения любой двигательной задачи
привлекается относительно большое число. мышц или мышечных групп.
Базовый потенциал силы, зависящий главным образом от поперечного
сечения мышечного волокна, объема мышцы, строения волокон и внутримышечной
координации, может быть превращен в оптимальный результат только
в том случае, когда отдельные мышцы или мышечные группы будут
задействованы в пространственно-временном и динамико-временном
отношениях последовательно в соотвйствии с двигательной задачей.
Это также означает, что нервная система настраивается на использование
только тех мышц, работа которых необходима для решения определенной
двигательной задачи. Это взаимодействие участвующих в движении
мышц или мышечных групп называется межмышечной координацией.
Она всегда связана с определенным видом движения и не может
переноситься с одного движения на другое. Например, при выполнении
жима лежа на скамейке участвуют одни мышечные группы, а при подтягивании
на перекладине - другие. Взаимодействие мышц также организовано
по-разному. Особое значение для межмышечной координации имеет
согласованность в работе мышц, реализующих определенное движение
(агонисты) и мышц, действующих в противоположную сторону при этом
движении (антагонисты).
При скоростно-силовом жиме лежа на скамейке трицепс разгибает
руку в локтевом суставе, а бицепс, плечевая мышца и плечелучевая
мышца (антагонисты), основная функция которых состоит в сгибании
руки в локтевом суставе, препятствуют разгибанию, особенно в конечной
фазе движения (см. табл. 13 и рис. 60 а). Взаимодействие агонистов
и антагонистов происходит следующим образом (упрощенно). При скоростно-силовом
выпрямлении руки из положения лежа на спине, преодолевая относительно
высокое сопротивление, одновременно серией коротких импульсов
активизируется максимально возможное количество двигательных единиц
трицепса и мышц, работающих вместе с ним (например, большая грудная
мышца и др.). За серией импульсов и начинающимся сокращением следует
сигнальная блокировка (биоэлектрическое молчание), во время которой
мышечные волокна укорачиваются с высокой скоростью без какого-либо
управления.
Сигнальная блокировка в двигательных единицах, реализующих движение
(трицепс и др.), связана с сигнальной блокировкой двигательных
единиц антагонистов, поэтому движение может выполняться беспрепятственно
и без потери силы. В конце движения для его торможения начинают
активизироваться антагонисты. Ариель (1976-1977) излагает этот
метод работы нервно-мышечной системы следующим образом. При обычном
подъеме штанги происходит начальный взрыв мышечной активности
в связи с тем, что мышца-агонист сокращается, а мышца-антагонист
расслабляется. Это приводит к ускорению движения конечности. Затем
следует спокойный промежуточный период в это время в связи с сокращением
антагониста происходит замедление движения конечности. В конце
движения напряжение мышцы-антагониста должно его остановить (7).
Эта согласованность в работе агонистов и антагонистов во многом
зависит от растянутости мышц. Преимущество растянутых мышц
в том, что они в состоянии покоя слегка напряжены (около 15% своей
длины равновесия) и из этого начального состояния способны развить
особенно большую силу. С другой стороны, они позволяют производить
движения с большой амплитудой, благодаря чему используется более
длинный путь ускорения и имеющийся потенциал силы. Большая амплитуда
позволяет выполнять движения мягче, эластичнее и плавнее, так
как антагонисты начинают их притормаживать позднее (см. 2.7.).
Чем больше мышц или мышечных групп принимают участие в движении,
т.е. чем сложнее движение, тем большую роль играет межмышечная
координация для выполнения силового упражнения.
В тренировочной практике различных видов спорта сложные технические
действия часто расчленяются на отдельные части (элементы). Цель
такого расчленения состоит в том, чтобы силовой тренировкой развить
мышечные группы, которые принимают непосредственное участие в
выполнении соревновательного движения. Усиление этих мышечных
групп осуществляется специальными упражнениями, структура которых
совпадает лишь с частью структуры соревновательного упражнения.
С помощью несложных специальных упражнений можно особенно сильно
нагружать мышцы, участвующие в соревновательном движении, и таким
образом эффективнее их развивать.
Такими специальными упражнениями, например, для толкателя ядра
являются жим лежа на скамейке, наклоны туловища в стороны, поднимание
и опускание туловища, приседания и вставание на носки. Новое,
хорошо скоординированное при помощи специальных упражнений взаимодействие
мышечных групп, укреплявшихся изолированно, требует специального
обучения технике, присущей данному виду спорта. Трудности проявляются
тогда, когда не все мышцы развиты достаточно хорошо, нарушена
гармония в развитии. Если, например, толкатель ядра при помощи
специальных упражнений улучшает лишь силу мышц-разгибателей рук
и ног и выпустит из внимания мышцы туловища, то это может привести
к серьезным нарушениям межмышечной координации. Финальное усилие
при этом останется небольшим. Если межмышечная координация не
удовлетворяет предъявляемым требованиям, например, когда при взрывном
включении мышц начальная иннервация агониста не связана с оптимальным
биоэлектрическим молчанием (компенсация потерь) антагониста, или
когда его силы торможения включаются преждевременно, то теряется
большая часть проявляемой силы. В отличие от этого высокий уровень
межмышечной координации проявляется в оптимальной плавности движения,
целесообразном ритме, точном выполнении и, в конечном итоге, в
большой силовой отдаче.
За счет целенаправленной тренировки, которую часто называют также
тренировкой на развитие техники с помощью силовых элементов, можно
значительно поднять уровень межмышечной координации. Однако следует
сказать еще раз, что взаимодействие мышц, как правило, улучшается
лишь в натренированном движении. На движение с похожими пространственно-временными
и динамико-временными структурами это улучшение переносится лишь
частично, а на движения с неодинаковыми структурами не переносится
совсем. Высокий базовый потенциал силы является основной предпосылкой,
но никак не гарантией того, что движения будут выполняться с полной
силой. Лишь тот спортсмен способен целесообразно использовать
свой потенциал силы, который понял, что отработка межмышечной
координации должна проводиться на соответствующих движениях. Тренированный
спортсмен, в отличие 'от нетренированного, обладает не только
большей мышечной массой и может ее гораздо полнее использовать
(внутримышечная координация), он также в состоянии гораздо эффективнее
реализовать этот потенциал силы в своем спортивном результате
(межмышечная координация).
2.2.3. Энергетическое обеспечение мышечной деятельности
2.2.3.1. Энергоносители
Для выполнения работы мышца должна быть обеспечена энергией. Мышечными
энергоносителями являются, главным образом, фосфатные соединения,
обладающие большими энергетическими запасами (аденозинтрифосфат,
креатинфосфат), углеводы (глюкоза, гликоген) и жиры. Белки как
энергоносители играют второстепенную роль, однако для увеличения
объема, роста мышцы они имеют первостепенное значение. Лишь в
редких случаях (голодание, предельные и продолжительные нагрузки)
белки в виде аминокислот могут принимать участие в энергетическом
обмене веществ.
В мышце накапливаются богатые энергией фосфаты соединения, гликоген
и жиры. Гликоген и жиры откладываются также в печени и в подкожной
жировой клетчатке. В человеке на 1кг мышечной массы приходится
от 3,5 до 7,5 ммол аденозинтрифосфата (АТФ) и 16-28 ммол креатин-фосфата
(КФ). Это соответствует собственным энергетическим резервам организма
в АТФ - около 5 кдж (1,2 ккал) и в КФ - приблизительно 15 кдж
(3,6 ккал). Энергетические резервы, хранящиеся в организме в форме
гликогена, составляют у нетренированного человека около 7 500
кдж (1 800 ккал, 450 г), у тренированного -до 13 000 кдж (31 ккал,
750 г). Из 'этого количества на гликоген печени приходится около
2 600 кдж (620 ккал, 150 г). Мышечный гликоген представляет собой
энергетический резерв, быстро включающийся в знергообразование.
И в энергетическом отношении он эффективней, так как его не надо
транспортировать к работающей мышце сначала по кровеносному руслу,
а затем, как через шлюз, пропускать через оболочку клетки.
Далее следует учитывать тот факт, что мышечное волокно относительно
легко принимает подаваемую ей по кровеносному руслу глюкозу и
накапливает ее в форме гликогена, но очень неохотно, лишь в небольших
количествах, возвращает в кровоток гликоген для потребления его
другими, интенсивно работающими мышцами. Кроме того, гликоген
печени лишь частично может быть использован для мышечной деятельности,
так как жизненно важные функции головного мозга также должны постоянно
обеспечиваться питанием глюкозой, доставляемой по кровеносным
сосудам. По этой причине разнообразные защитные механизмы препятствуют
чрезмерному потреблению гликогена печени и тем самым снижению
уровня сахара в крови, довольно постоянному в нормальных условиях
(80-90 мг глюкозы на 100 мл крови).
Жировой запас, насчитывающий от 125 000 до более чем 400 000 кдж
(от 30 000 до более чем 100 000 ккал), является практически неиссякаемым
источником энергии при выполнении продолжительной спортивной работы
небольшой интенсивности. Наибольшая доля згих запасов содержится
в подкожной жировой клетчатке; мышечные жировые тела (капли триглицерида),
быстрее включающиеся в работу, накапливают лишь около 8 000 кдж
(1 900 ккал).
2.2.3.2. Анаэробное энергообразование
Непосредственным источником энергии для мышечных волокон всегда
является аденозинтрифосфат (АТФ). Он расщепляется на аденозиндифосфат
(АДФ) и фосфат (Ф). При расщеплении выделяется энергия (38-42
кдж = 9-10 ккал/на мол. расщепленного фосфата).
АТФ->АДФ + Ф + Энергия
Лишь около трети этой высвобождающейся энергии превращается в
механическую работу; большая часть выделяется в виде тепла. В
связи с тем, что содержание аденозинтрифос-фата в мышце довольно
ограничено (3,5-7,5 ммол/кг), уже через очень короткое время предельно
интенсивной работы (1-3 с) его мышечные запасы иссякают. И если
бы целая система различных, частично одновременно протекающих
процессов, не контролировала бы и не восстанавливала аденозинтрифосфат,
т. е. не снабжала бы мыппцы энергией, то работу пришлось бы прекратить.
Восстановление (ресинтез) АТФ осуществляется при помощи относительно
простого биохимического процесса. Возникающие в результате расщепления
АТФ продукты АДФ и Ф соединяются снова.
АДФ + Ф + Энергия -> АТФ
Для этой биохимической реакции, которую иначе называют фосфорилированием,
требуется энергия. Она получается за счет расщепления других энергоносителей.
В какой форме и с помощью каких энергоносителей будет осуществлено
восстановление аденозинтрифосфата, зависит от энергии, требуемой
на единицу времени. При внезапной, осуществляемой из состояния
покоя, очень интенсивной мышечной работе АТФ восстанавливается
с помощью креатинфосфата
КФ + АДФ -> Креатин (К) + АТФ
Так как один моль креатинфосфата производит приблизительно один
моль АТФ, то выход энергии в результате этого процесса соответствует,
примерно, энергии, получаемой от расщепления АТФ.
Креатинфосфат содержится в мышце также в незначительных количествах
(16-28 ммол/кг). Несмотря на то, что имеющимся количеством креатинфосфата
можно образовать в три раза больше энергии, чем при помощи АТФ,
эти запасы будут сильно истощены через 7-12 с предельно интенсивной
работы и через 15-30 с интенсивной работы. Работу необходимо или
прервать, или же продолжать с меньшей интенсивностью. Для продолжения
работы с меньшей интенсивностью требуются другие энергоносители.
Гликоген, содержащийся в мышце в таких условиях расщепляется без
участия кислорода на молочную кислоту (лактат), обладающую меньшей
энергоемкостью, чем АТФ.
Гликоген -> Лактат + АТФ
При этом один моль мышечного гликогена поставляет 3 моля АТФ.
Наибольшей активности этот биохимический процесс достигает через
несколько секунд интенсивной работы. Перед этим должна быть снижена
до соответствующей величины концентрация более эффективных в энергетическом
отношении фосфатов. Восстановление АТФ путем расщепления гликогена
(при отсутствии кислорода) высвобождает за одну и ту же единицу
времени лишь около трети энергии, которую можно было бы получить
расщеплением фосфата. Поэтому интенсивность работы необходимо
уменьшить. Эти процессы - причина того, что, например, бегун на
100-метровую дистанцию, пробежав около 80 м, часто теряет скорость:
его запасы фосфатов, богатых энергий, сильно исчерпаны, и организм
вынужден переключиться на получение энергии из менее эффективного
гликогена.
При выполнении отдельных упражнений и коротких серий упражнений
(3-30 с) в тренировке максимальной и скоростной силы энергия для
мышечного сокращения также выделяется со „складов" АТФ и
КФ. Для серий интенсивных упражнений продолжительностью 10-30
с энергия получается в большей степени за счёт использования гликогена.
На рис. 10 изображены процессы выделения энергии при интенсивной
спортивной работе в зависимости от длительности нагрузки.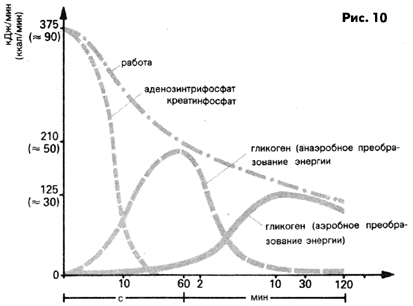
Рис. 10 Преобразование энергии (кДж/мин) в скелетной мускулатуре
человека во время спортивной работы в зависимости от длительности
нагрузки (преобразовано по Ховальду)
Мышечная работа длительностью около 2 мин, при выполнении которой
энергию получают изложенным выше способом, осуществляется преимущественно
быстрыми FT-волокнами, накапливающими энергоемкие фосфатные соединения
и гликоген в относительно больших количествах. Как уже отмечалось,
при расщеплении гликогена образуется лактат. Кислые продукты обмена
веществ (лактат, СО2 и т.д.), производимые очень быстро, в процессе
интенсивной работы не могут компенсироваться, быстро разлагаться
или выделяться ни при помощи имеющихся в крови буферных веществ,
ни за счет дыхания (СО2. После нагрузок, приводящих к сильному
утомлению, концентрация лактата в мышце поднимается до ЗОммол/л
и в крови до 20 ммол/л. После выполнения работы „до отказа",
т.е. работы, во время которой интенсивно участвует большое количество
мышечной массы, затрудняется дыхание и сдерживается приток и отток
крови в пределах до 25-28 ммол/л (например, после интенсивно проведенной
борцовской схватки).
Большое количество лактата в значительной степени способствует
переокислению организма, сковывающему мышечную деятельность. Увеличивающееся
закисление организма затрудняет все в большей степени дальнейшее
расщепление гликогена и, как следствие, препятствует ресинтезу
АТФ. Однако, энергия АТФ нужна не только для сокращения, но и
для расслабления мышцы (разрыв актино-миозиновых мостиков). При
нехватке АТФ существенно ограничивается его так называемое „смягчающее"
действие, которое к тому же имеет большое значение для растягиваемости
мышцы. Мышца поэтому расслабляется с каждым разом медленнее и
в конце концов ее сводит судорогой. Это происходит несмотря на
то, что для нагрузок подобной интенсивности часто затрачивается
лишь треть всех запасов мышечного гликогена. В практике иногда
говорят о „скисании" спортсмена. Работу следует остановить,
если своевременно не будет подключена другая форма получения энергии,
которая в свою очередь заставит снизить интенсивность нагрузки
на единицу времени. При использовании методов больших нагрузок
для развития силовой выносливости (например, интенсивный интервальный
метод, см. 9.2.3.) фазы интенсивной работы следует чередовать
с короткими паузами отдыха. Во время этих интервалов отдыха кислые
продукты обмена веществ могут 'частично удаляться из мышечных
волокон, а влияние других факторов, ведущих к утомлению, может
быть ослаблено. Благодаря этому помехи при расщеплении гликогена
и восстановлении АТФ будут не столь значительными, и спортсмен
через относительно небольшой промежуток времени может вновь приступить
к работе. Однако слишком короткие паузы препятствуют восполнению
запасов гликогена. Из-за этого с каждой дозой нагрузки их становится
все меньше и меньше.
Интервальные нагрузки приводят к гораздо более сильному опустошению
гликогеновых депо, чем однократные интенсивные нагрузки. Это следует
иметь в виду не только в период силовой тренировки, но и при занятиях
видами спорта со сменяющимися по интенсивности нагрузками (футбол,
гандбол, борьба и т.д.). Изложенные биохимические процессы протекают
практически без участия кислорода. Поэтому их называют анаэробными
процессами. Расщепление богатых энергией фосфатов именуется
анаэробным алактатным (проходящим без образования лактата) процессом,
а разложение гликогена называется анаэробным лактатным (проходящим
с образованием лактата) процессом.
2.2.3.3. Аэробное энергообразование
Гликоген может распадаться не только на молочную кислоту (лактат),
имеется также возможность окислить гликоген при участии кислорода
(О2). При этом наряду с энергией выделяются вода (Н2О) и углекислый
газ (СО2).
Гликоген + О2 -> Н2О + СО2 + АТФ
Этот процесс сгорания углеводов при участии кислорода называется
аэробным путем получения энергии. При этом один моль глюкозы (при
разложении мышечного гликогена) поставляет 39 молей АТФ. Окисление
гликогена с участием кислорода почти в 13 раз эффективнее, чем
его расщепление без кислорода.
При интенсивных нагрузках продолжительностью около 5 мин 50% энергии
производится посредством анаэробного и 50%-посредством аэробного
обмена веществ. Если длительность интенсивной нагрузки менее 5
мин, то в этом случае большее значение приобретают анаэробные
процессы; если нагрузка продолжается более 5 мин, то в преобразовании
энергии неизбежно повышается доля аэробного обмена веществ. Относительно
высокая доля анаэробных процессов приводит к высокому содержанию
лактата в крови (15-25 ммоль/л). В этих условиях мышце начинает
недоставать своих собственных энергетических ресурсов. Гликоген
печени в виде глюкозы с кровью доставляется к мышце и способствует
покрытию энергетического дефицита.
Наряду с FTO-волокнами в работе продолжительностью 2-10 мин, примеры
которой здесь уже приводились, широко принимают участие также
медленные ST-волокна (красные волокна). По сравнению с быстрыми
РТ-волокнами в FTO и особенно в ST-волокнах содержится больше
миоглобина. Миоглобин связывает, „выбирает" кислород, а затем
предоставляет его в распоряжение митохондрий. Кроме того, в этих
волокнах содержится больше крупных митохондрий, в которых протекают
аэробные процессы обмена веществ. По этим, а также другим причинам
ST-волокна и FTO-волокна наиболее пригодны для аэробного преобразования
энергии и тем самым для выполнения работы, требующей выносливости.
При усиленных нагрузках продолжительностью свыше 10 мин доминирует,
без сомнения, аэробный обмен веществ, посредством которого получают
70-95 % требуемой энергии. Эта продолжительная работа с приложением
силы менее 25% от максимальной реализуется в первую очередь ST-волокнами.
Анаэробные процессы обмена веществ и FT-волокна включаются в движение
при выполнении этой продолжительной работы прежде всего в начальной
фазе, а также для преодоления чередующихся внешних сопротивлений
(например, на промежуточных рывках или подъемах в горку в лыжных
гонках и велоспорте). В связи с незначительной долей анаэробного
пути получения энергии при долговременных нагрузках накопление
лактата в мышце и крови остается незначительным (3-14 ммол/л в
крови). С увеличением продолжительности нагрузок на передний план
выходят жиры, являясь источником аэробного преобразования энергии.
Их распад происходит в принципе так же, как и аэробное разложение
гликогена в конечные продукты: воду и углекислый газ. Жиры могут
накапливаться в мышечном волокне в виде маленьких капелек (капли
триглицерида) или в виде жирных кислот могут транспортироваться
по кровеносному руслу к работающей мышце из подкожной жировой
клетчатки. Однако следует учитывать, что интенсивность нагрузки
постоянно возрастает, в том числе и в тех спортивных дисциплинах,
где требуется чрезвычайно высокая выносливость.
Энергетическое обеспечение зтой относительно интенсивной мышечной
деятельности осуществляется большей частью, несмотря на участие
жиров, за счет распада резервов гликогена (мышечный и печеночный
гликоген). Так, например, доля жиров в производстве энергии у
марафонца мирового класса составляет всего около 20%.
При усиленных нагрузках продолжительностью свыше 90 мин. собственных
резервов (гликоген) организма для продолжения работы часто не
хватает. Поэтому эти резервы должны пополняться дополнительными
питательными веществами (напитки с богатым содержанием глюкозы
и минеральных веществ).
Интересно к прочтению:
| Аминокислоты. |
 |
Раньше о таких вещества как аминокислоты многие люди, в том числе и те, кто на сегодняшний день являются заядлыми спортсменами, знали лишь из школьного курса химии и биологии. |
| Серджио Олива |
 |
Самым первым чернокожим получившим звание «Мистер Олимпия» стал выходец с Кубы Серджио Олива. Родившись в Гаване четвертого июля 1941-ого года, Серджио после 1959 года, когда Фидель Кастро взял власть в стране, всеми способами хотел сбежать с социалистической Кубы. |
| Ларри Скотт |
 |
Ларри Скотт появился на свет в Соединенных Штатах Америки (штат Айдахо) в октябре 1938 года. Сегодня это может показаться несколько странным, но и в детстве, и в юном возрасте будущий спортсмен совершенно не отличался мускулистостью. |